Our lab has 6 presentations at this year’s Annual Meeting of the Society for Neuroscience (SfN) in Chicago, IL. Our presentations cover topics related to perception, action, and neurophysiology, as part of our long term effort to better understand the physiology of active perception in the brain.
1. The influence of visual forms on saccades
Amar’s presentation demonstrates that peripheral objects confer an advantage to saccadic eye movements, in a manner that appears to “capture” looking at meaningful visual forms (as opposed to noise or abstract spots). This means that the oculomotor system has privileged access to peripheral object detection processes. This is an important finding because a lot of work on object recognition focuses on objects being already presented at the line of sight. However, for us to interact with our environment, we first need to make saccades to align our gaze with objects of interest. Amar’s work shows that the oculomotor system benefits from peripheral object detection mechanisms to facilitate orienting. This work is in line with our findings that oculomotor brain structures like the superior colliculus prioritize visual processing for the advantage of orienting behaviors (e.g. our 2018, 2018 papers or our 2016 paper).
2. The impeccable accuracy of saccades
Continuing with saccades, Tanya’s presentation shows that saccades are much more accurate than is generally assumed. Very often, people argue that saccades are sloppy: that they bring the line of sight rapidly to “approximately” the region of interest where the object lands in the fovea. Then, according to the argument, as long as the image is in the fovea, then the saccades have achieved their purpose. However, foveal processing itself is not uniform (e.g. our 2019 paper). Moreover, it is not entirely known with which “resolution” saccades can resolve tiny position displacements of peripheral targets. So, Tanya designed experiments in which humans and monkeys were making horizontal (rightward or leftward) or vertical (upward or downward) eye movements. She displaced the target orthogonally by a very tiny amount. For example, if the saccade was to the right, then on some trials, the target was displaced ever so slightly upward or downward from “pure right”. She found that saccades curve in very systematic ways to exactly intercept the shifted target. She also related these corrections to perceptual judgements in humans. This work is important because it demonstrates that even if gaze is variable at fixation, which results in small shifts in the retinotopic image of saccade targets, the oculomotor system has sufficient accuracy “resolution” to account for such variability. This work is related to our 2016 findings of functional and structural discontinuities in the representation of rightward/leftward or upward/downward axes in the superior colliculus.
3. The origin of perceptual saccadic suppression
Matthias is also interested in saccades, but this time, focusing on their influence on visual perception. As is well known, saccades are very rapid and “ballistic” eye movements. They thus cause a huge motion flow of images on the retina as the eye rotates at high speed. If a brief visual flash is presented around such rapid motion flow, its perceptual detectability is reduced dramatically. This phenomenon is known as saccadic suppression. One intriguing property of saccadic suppression is that if the brief visual flash has different properties, it experiences different “amounts” of suppression. Specifically, coarse-type flashes are suppressed strongly, whereas fine image details are not. This is a robust perceptual property of saccadic suppression that was known for 25 years. Its mechanisms, on the other hand, are completely missing (but see our 2017 paper). The general assumption is that this “selective” property of strong coarse suppression and weak fine suppression is emerging through an internal motor command causing “selective” suppression. However, Matthias found that such suppression does not even need a saccade at all to occur. If one simulates the motion flow of saccades on the retina, then the same suppression happens. Moreover, with simple image tricks, one can get completely rid of “selective” suppression in the first place, even with real saccades. Therefore, with or without saccades, selective suppression may or may not happen depending on the visual flow on the retina. This work is important because it recasts the interpretation of a popular perceptual phenomenon. It is related to our recent work, like in 2010, 2015, 2017, and 2018.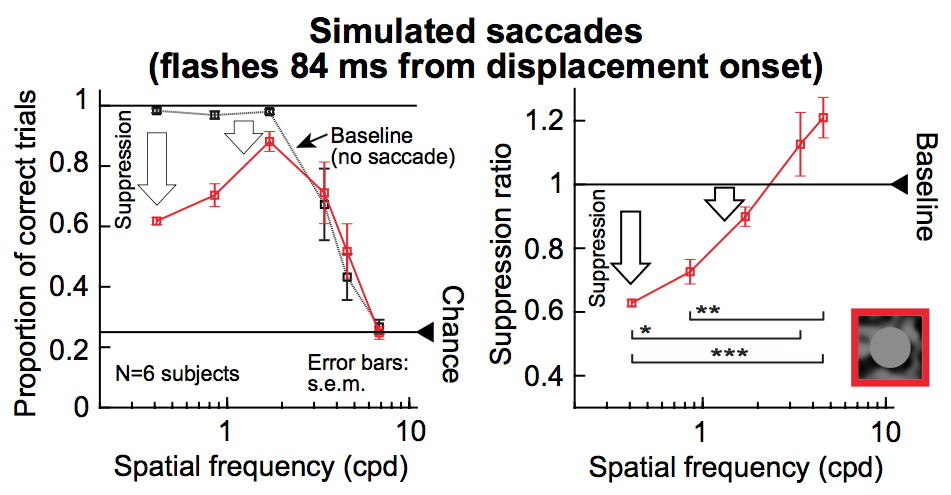
4. The retinal mechanisms of perceptual saccadic suppression
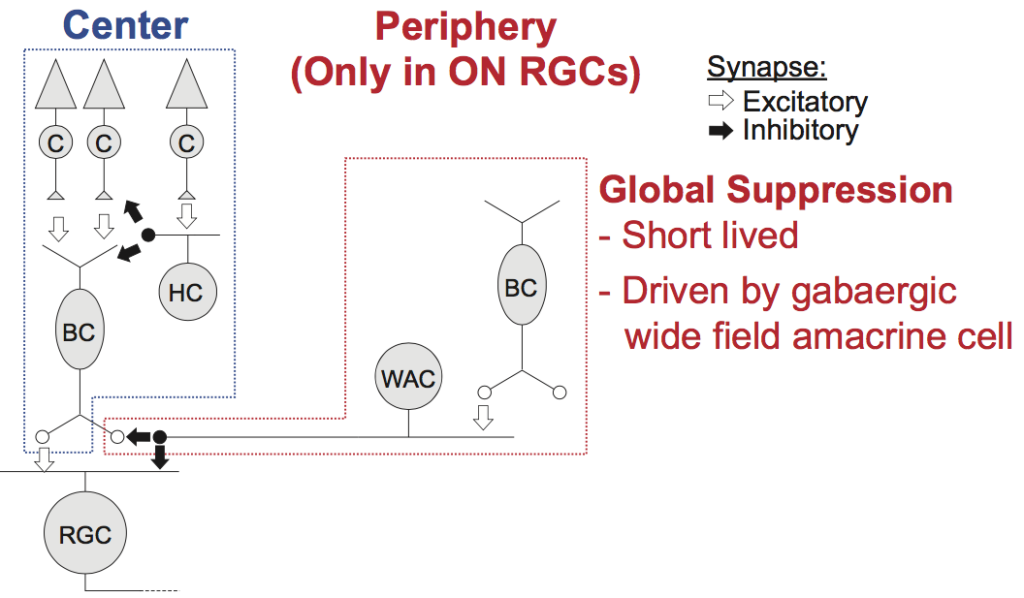
Related to Matthias’ work, Saad has been exploring retinal-circuit mechanisms of the impacts of saccade-induced visual flow and brief flashes. He finds that a combination of local and global image processing mechanisms mediate the starting point of perceptual saccadic suppression. Coupled with Matthias’ work above, this work is important because it provides a decidedly fresh perspective on the neural mechanisms and origins of a highly robust perceptual phenomenon, saccadic suppression, that has intrigued philosophers and scientists for many decades.
5. The importance of every single superior colliculus spike for saccade generation
Antimo’s work now fills the gap between behavior and input (retinal processing). He is exploring neural activity in the superior colliculus, a brain area that is both an early visual area (because it receives early visual information, including directly from the retina) as well as a late motor area (because it influences saccade generation). What Antimo did was to artificially “inject” action potentials in the superior colliculus at the time of saccade generation. He did so either with visual stimuli that appeared suddenly or with electrical microstimulation in the superior colliculus. With careful analysis, he found that every single action potential of a given neuron matters for the metrics of saccades. He specifically could alter the size of the planned saccade in a very lawful and predictable manner by the number of action potentials that he has “injected” into the superior colliculus. This work is important because there is continued discussion on what “code” the superior colliculus uses to drive saccadic eye movements. The work is also important because it addresses an important topic, which is perhaps less studied as it should, on the interplay between ongoing motor behavior and all sources of interruptions that might happen from the world. The work is related to our previous results from 2013 and 2017.

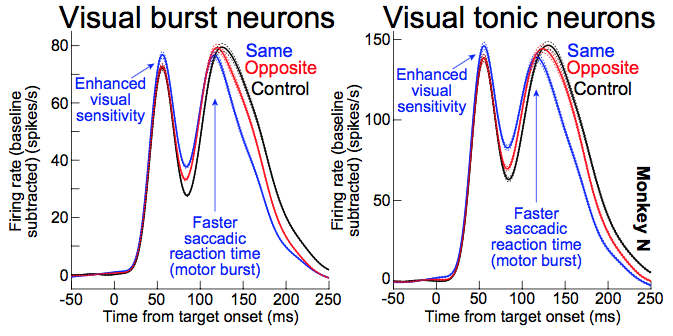
6. The role of foveal motor programming of microsaccades in influencing peripheral visual performance (such as in covert visual attention)
Continuing with the superior colliculus, Tian used our retinal image stabilization technique (e.g. 2013, 2016, 2018) to causally control foveal motor error. He found that this is sufficient to completely bias microsaccade directions. He then recorded superior colliculus neurons in extra-foveal sites, and he found that there was enhanced visual sensitivity and faster behavioral performance whenever microsaccades were biased in a certain direction. This means that the generation of “foveal action” in the form of microsaccades is causally sufficient to modulate peripheral visual sensitivity and behavioral performance in the same direction as the microsaccades. These effects mirror exactly our earlier findings (2015, 2013), but they now show causal evidence for the role of microsaccades. This is important because our extensive work on the links between microsaccades and peripheral covert visual attention (e.g. 2002, 2011, 2013, 2013, 2015) has often been met with resistance from the attention community. One argument that was put forward by the opponents of our work was that microsaccades are just a “dirty readout” of the state of peripheral brain sensitization (if the brain is sensitized because of attention, then the microsaccades might happen in a biased direction but still in a stochastic manner). Our work, however, shows that this is not necessarily the case. Microsaccades happen for very clear reasons and are not necessarily stochastic in this sense. Moreover, they cause modulations of peripheral activity, perhaps due to foveal magnification (2019). The current work put forth a strong test of our earlier findings: causally influence foveal motor error and see what happens to peripheral performance. The results supported our earlier suggestions. Moreover, Tong (our newest lab member) added to this presentation by performing perceptual performance experiments in naive human subjects, and found that perceptual performance of humans was also causally manipulated by controlling the generation of individual microsaccades. Therefore, not only superior colliculus extra-foveal visual sensitivity is causally influenced by microsaccade generation, but also perceptual performance in humans. These results are important because they suggest that the link between microsaccades and covert visual attention is much stronger than appreciated, especially by the covert attention community.