
We have a new paper now published in the Journal of Neurophysiology! In it, we continued our studies of visual interruption.
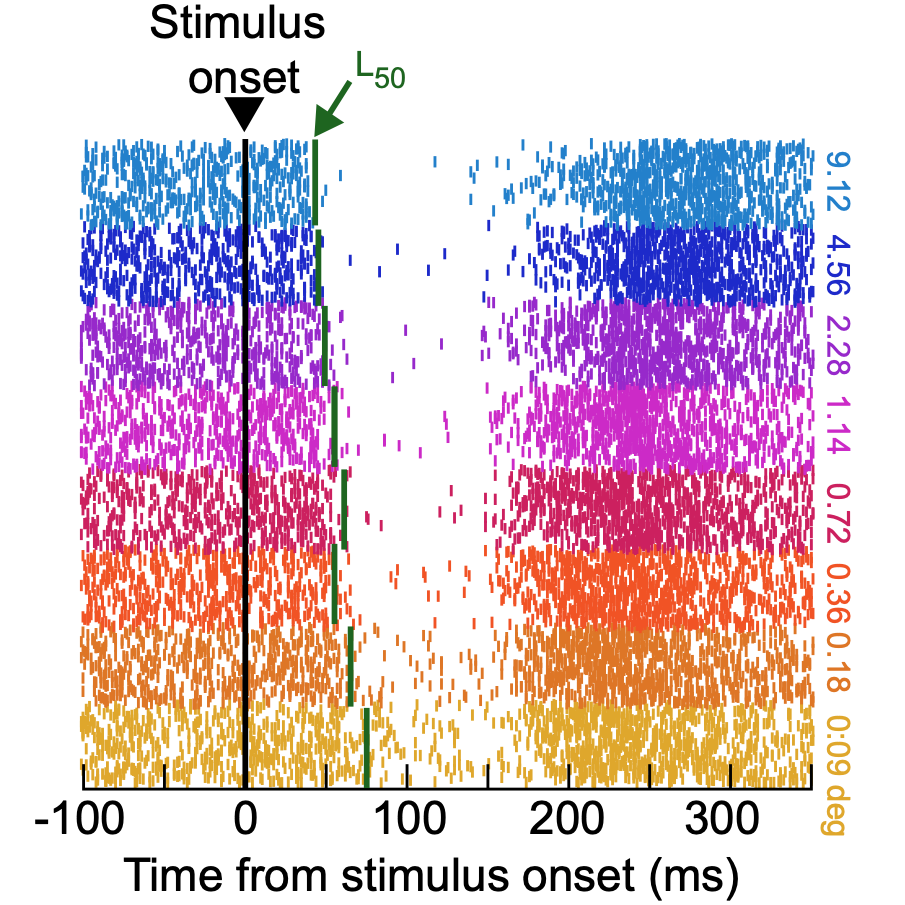
During natural visual behavior, we continuously generate saccadic eye movements. Whenever a visual transient occurs in the environment, the likelihood of saccade generation suddenly drops to almost zero within less than 100 ms from the stimulus event. We had recently written an exhaustive review article on how we think that this rapid visual interruption of saccade generation takes place, and why it is meaningful for the brain to behave in this manner. Critical in this phenomenon, and in our review, is the idea that visual signals impinging on the retina are able to very rapidly exert an influence on the final oculomotor generation pathways of the brain.
In this new work, we characterized the visual feature tuning properties of the interruption phenomenon. For example, might a low contrast stimulus cause saccadic inhibition earlier or later than a high contrast stimulus? Or, does the inhibition of saccades, instead, just have a constant short latency from stimulus onset, independent of the visual appearance of the stimulus? We tested several different feature dimensions, and we consistently observed that saccadic inhibition properties are robustly stimulus-dependent.
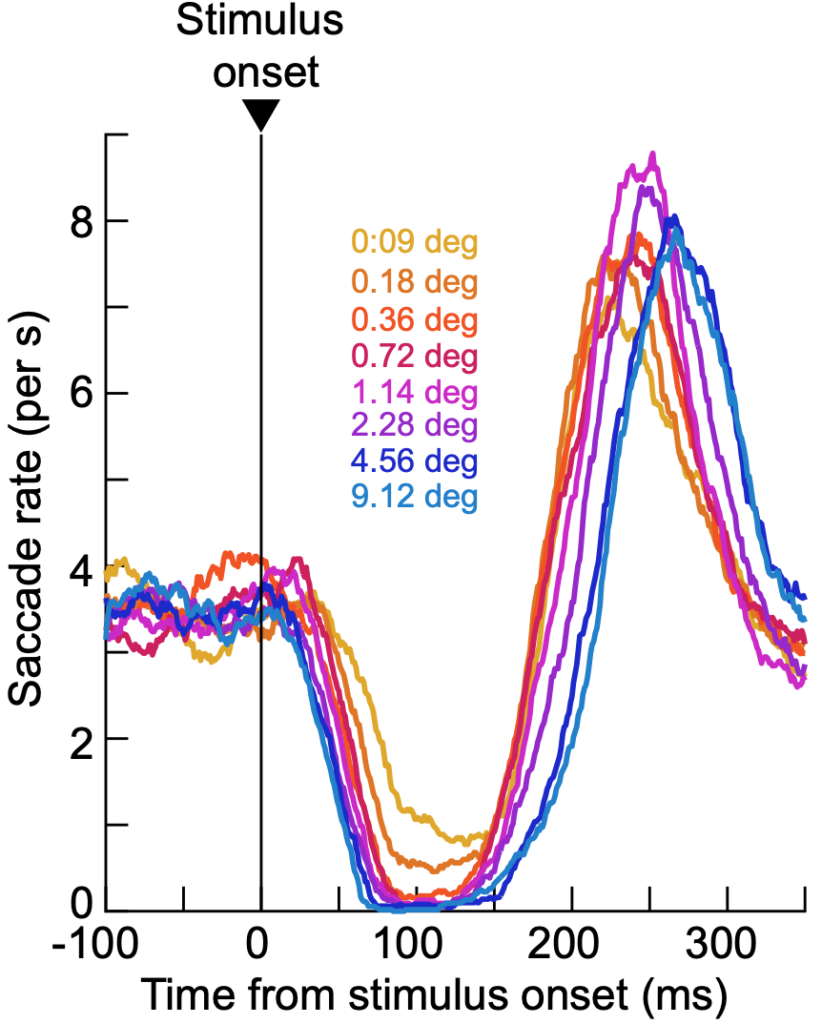
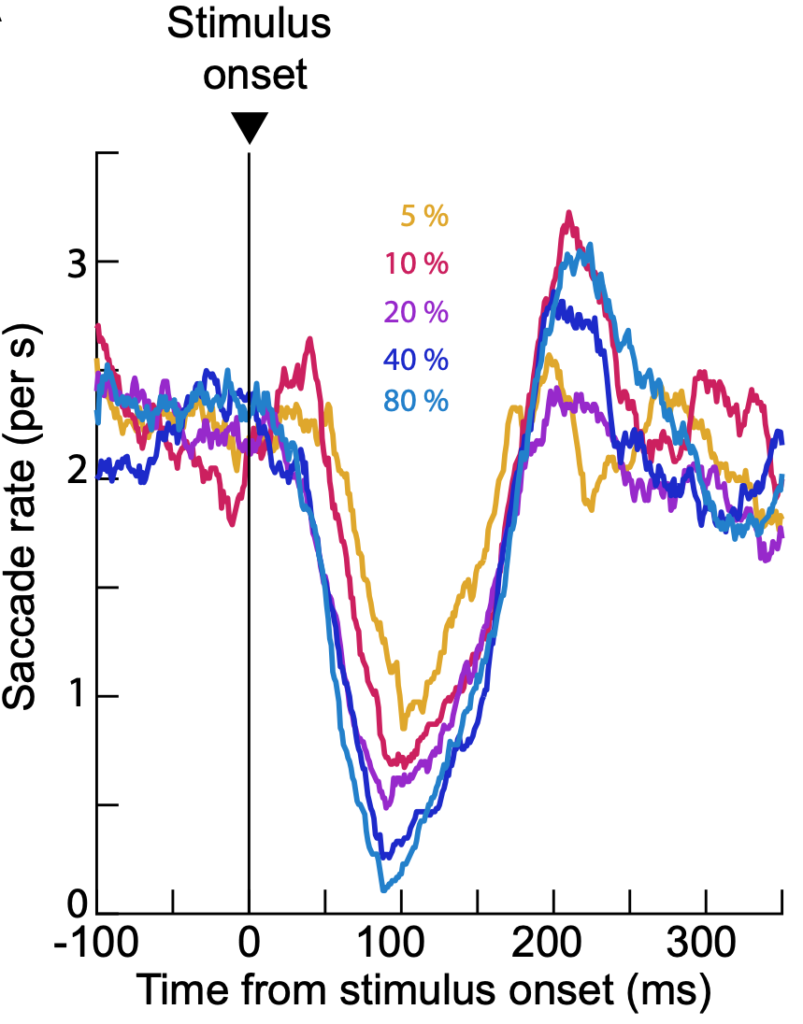
Interestingly, saccadic inhibition exhibited a kind of low-pass characteristic. Low spatial frequency stimuli cause earlier and stronger saccadic inhibition than high spatial frequency ones. This raises intriguing questions about the neurophysiology of saccadic inhibition. For example, while it might be the case that visual signals from the primary visual cortex (V1) ultimately reach the final oculomotor pathway to cause saccadic inhibition, these signals are reformatted by the oculomotor system: V1 might prefer band-pass stimuli, whereas saccadic inhibition is a low-pass phenomenon (like the superior colliculus). Thus, the lens with which saccadic inhibition “sees” the world is somewhat different from the lens with which perception might operate. This idea generates behind it interesting questions about the neural mechanisms underlying saccadic inhibition, and how signal flows, along with functional specializations in the visual-motor hierarchy, matter in sensory processing.
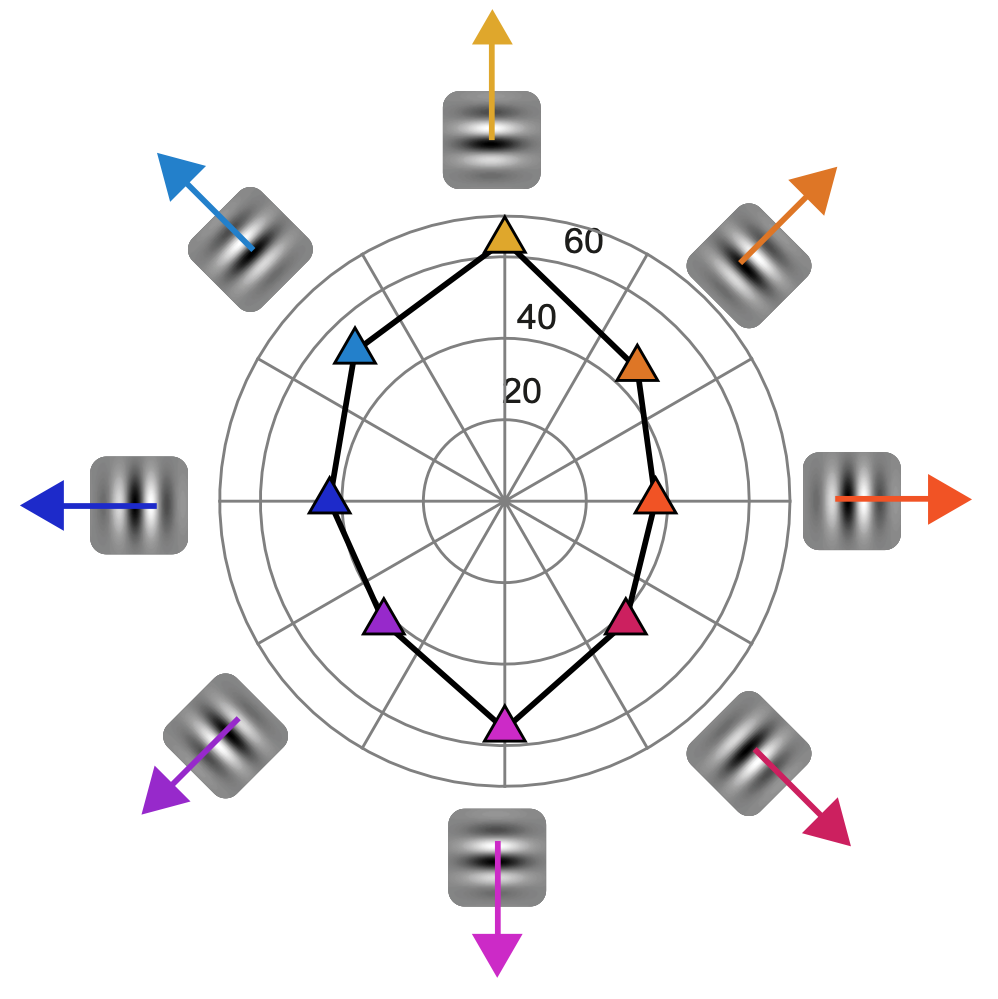
Besides spatial frequency and contrast, we also tested the influences of motion directions and orientation signals on saccadic inhibition. We observed interesting anisotropies in saccadic inhibition timing as a function of motion direction and orientation, which we did not initially anticipate. These anisotropies might be related to visual field anisotropies (as, for example, evidenced in our recent study here). We will definitely continue exploring this new observation in our future studies.
Stay tuned for future exciting neurophysiological observations building on this new latest paper from our lab!