

We have a new paper just published in eNeuro! In this paper, we developed a new, flexible behavioral paradigm for exploring peri-saccadic visual mislocalization in rhesus macaque monkeys.
Peri-saccadic visual mislocalization refers to the phenomenon in which your percept of a stimulus flash can be strongly altered by you making a rapid eye movement. In particular, if the flash is of high contrast and you can still detect it around the time of saccades (or rapid eye movements), then you tend to report seeing it at the wrong place (i.e. you grossly mislocalize it). This perceptual phenomenon is highly robust in humans, and we have studied it extensively in the past (see, for example, here and here). However, the underlying neurophysiological mechanisms for explaining it have remained elusive. This was due, at least in part, to the lack of a proper behavioral paradigm in monkeys, which constitute an ideal animal model for exploring the neurophysiological mechanisms of active vision.
In this latest work, we developed a robust peri-saccadic visual mislocalization paradigm for monkeys. The most important aspect of this paradigm is that it allowed for the use of arbitrary saccade directions, and also arbitrary peri-saccadic flash locations. This was very important for us to incorporate, especially since the neurons encountered during neurophysiological experiments (our ultimate goal) could have completely arbitrary locations.
We demonstrated that monkeys can be easily trained to report their peri-saccadic percepts, and we also demonstrated that we got similar mislocalization patterns to those observed in humans. Critically, we could have any saccade direction in the experiments, which, in addition to the above-mentioned constraints on neurophysiology, can also allow for exploring the asymmetries that might exist in peri-saccadic mislocalization patterns as a function of saccade direction. Indeed, in humans, we did find exactly such an asymmetry, which likely reflects strong asymmetries in the superior colliculus.
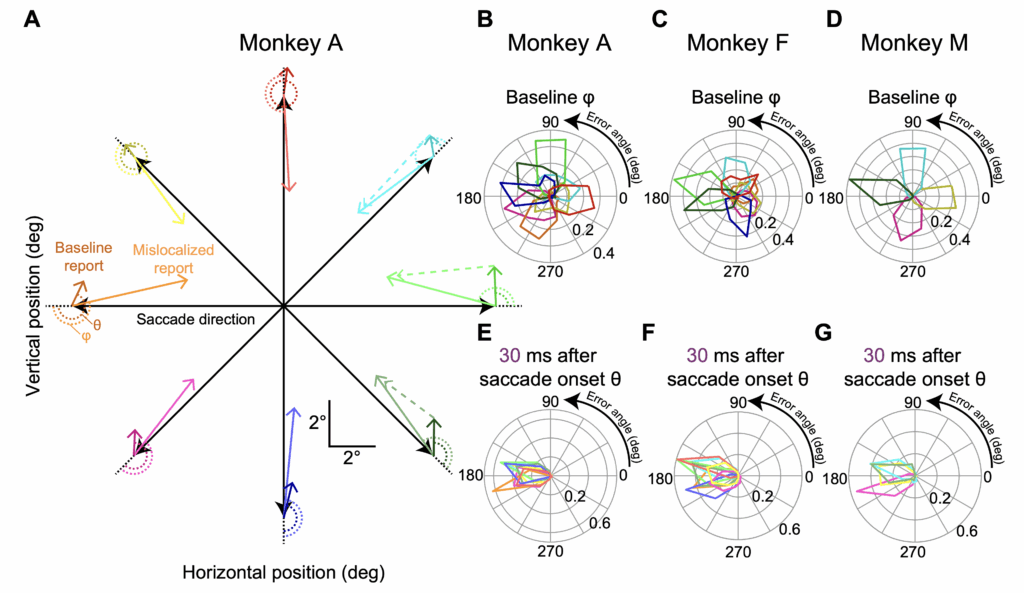
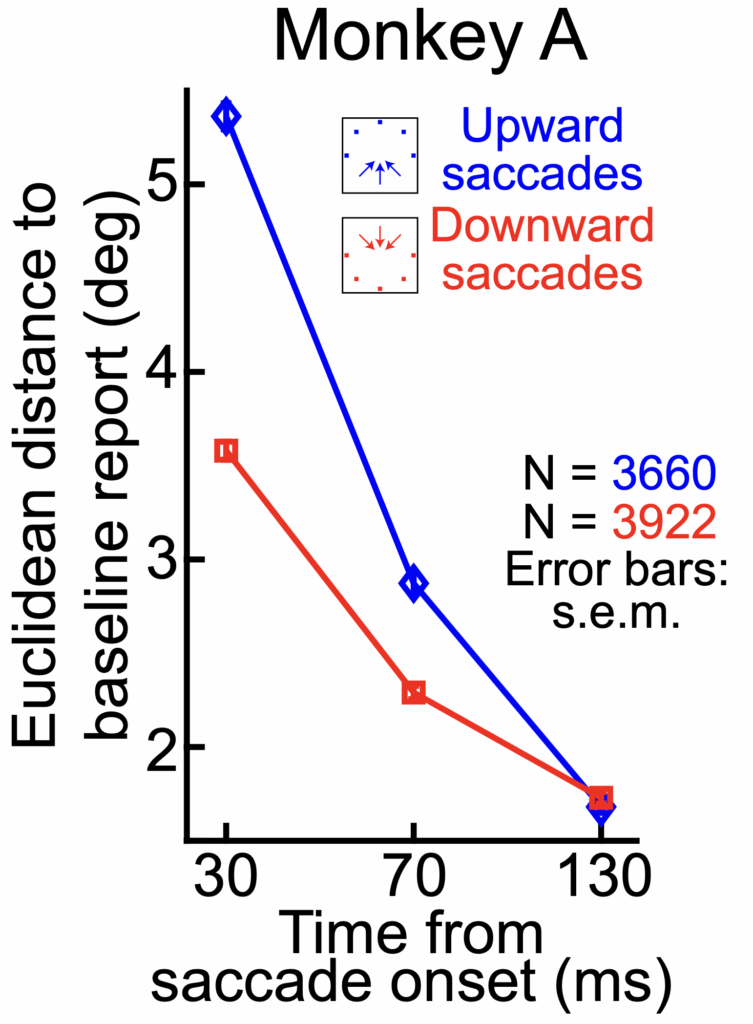
In fact, after demonstrating the basic phenomenon, we replicated our earlier human observations. For example, we found that mislocalization strength was much stronger for upward than downward saccades. Such replication is ideal to have, because it further justifies the use the monkeys for neurophysiological investigations.
The demo movie below shows how peri-saccadic mislocalization works. It shows two example trials. In the first trial (with green eye position traces), no mislocalization takes place. This is because the flash is presented for a very long time, including after the saccade. Specifically, the trial starts by directing your eye to the bottom left corner. This is where the monkey was fixating before generating the primary saccade. Then, the primary saccade is an upward-rightward saccade towards the screen center. At some point, a white square appears in the top of the display, and remains on for a very long time. After an experimentally enforced delay, the central saccade target disappears, and the monkey generates an upward saccade towards the recent location of the white square. That is, the monkey reports where it saw the flash by looking at its location. You can see in the demo movie that, for this trial, the report is relatively accurate. Then, in the second trial of the demo movie, the flash happens very briefly and only intra-saccadically (during the eye displacement from the bottom-left corner to the center of the display). After the enforced delay, the monkey has to report the flash location (with the second, report, saccade), and you can see that the monkey does so in a very erroneous manner. The final reported location (the end point of the report saccade) appears as if it was displaced backward in space along a direction directly opposite the upward-rightward direction of the original primary saccade. This is the essence of peri-saccadic visual mislocalization.
We look forward to plenty of very fruitful neurophysiological experiments with such a paradigm!