
Our lab participated in this year’s Vision Sciences Society Annual Conference, which took place in St. Pete Beach, Florida.

We had three presentations at this year’s meeting.
In her talk, Tanya presented more results on our intriguing observations that sound activates a dormant visual-motor pathway bypassing the primary visual cortex. This is a very interesting result because it demonstrates that multiple anatomical pathways from the retina to the eye movement output centers can be “recruited at will” depending on the task requirements! We’re currently extending these exciting results by inactivating the primary visual cortex while recording from the superior colliculus with and without sound stimuli.
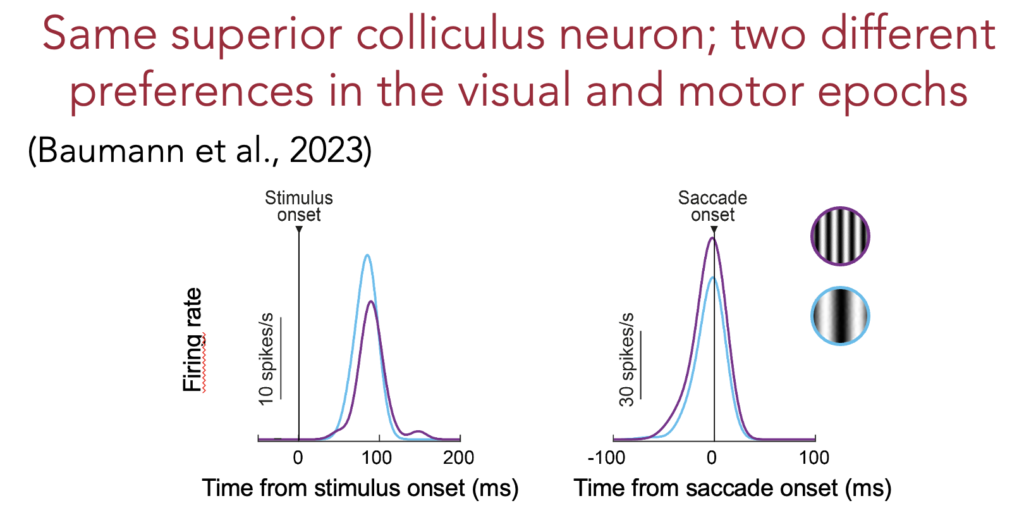
Next up, Tong presented brand new results being publicized for the very first time at an international conference. These results were concerned with what happens in the foveal visual representation of the superior colliculus when saccades are generated to peripheral targets. Normally, we and others always studied what the peripheral neurons did when generating saccades. However, our recent results and others made us wonder what happens in the fovea when saccades are about to be released. For example, a single peripheral neuron can prefer two different image features during a simple visual response epoch versus during saccade generation. If so, how can such a change in “mode” of the peripheral neuron between visual and motor regimes take place ?
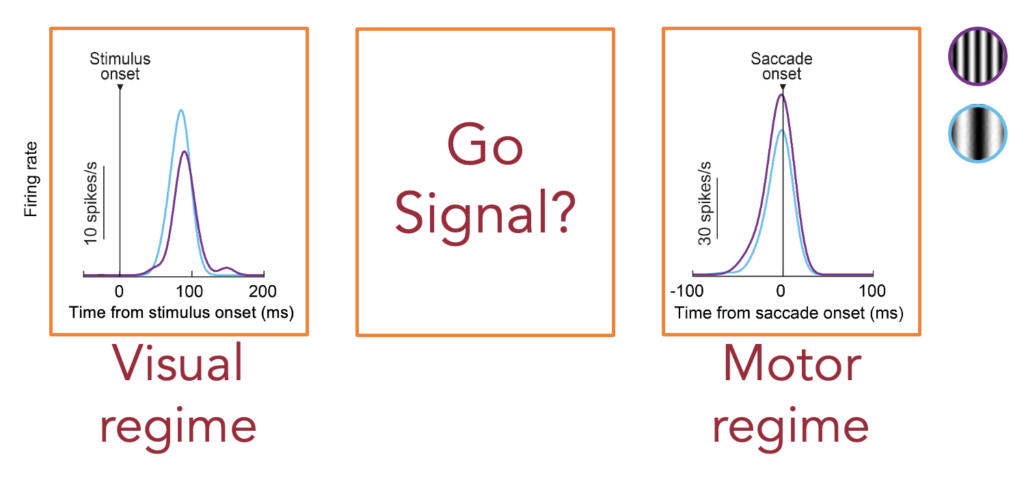
To answer this question, Tong decided to look at the time of the “go” signal instructing the subject to generate a saccade (i.e. instructing the subject to change mode from a visual regime during fixation to a saccade generation regime during eye movement command execution). What we found was that there was a transient change happening in the superior colliculus exactly during this state transition phase. Peripheral neurons representing the peripheral saccade target (like in the example neuron in the figure above) transiently completely paused their activity before they generated a strong motor burst. Remarkably, when we recorded foveal neurons, we found that they generated a transient burst instead!
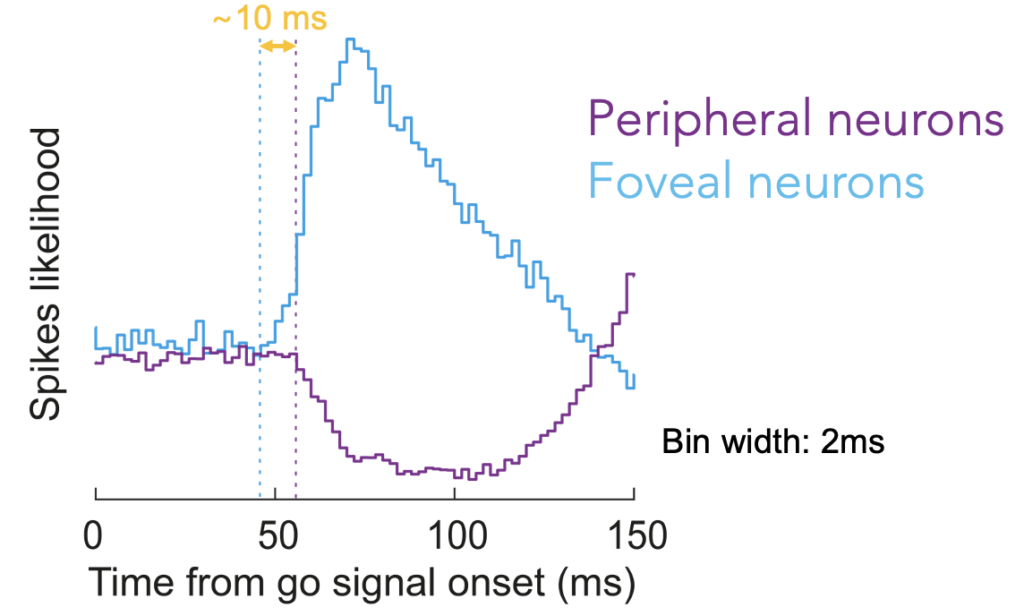
Perhaps more interestingly, the foveal neurons actually burst slightly earlier than when the peripheral neurons paused. Thus, we think that we found a foveal trigger signal for jumpstarting peripheral saccadic orienting! These results are interesting because they demonstrate that foveal processing is an active and integral component of eye movement generation even when the eye movements are concerned with peripheral targets outside the fovea.


Finally, Matthias continued our investigations of peri-saccadic suppression of visual sensitivity. He took our previous perceptual and retinal results, and he applied the same experiments in the superior colliculus and primary visual cortex. We found very interesting similarities and differences between the two brain areas. However, importantly, both areas replicated our previous retinal and perceptual findings. The differences between the two areas pointed to the possibility that the superior colliculus reflects perceptual effects better than the primary visual cortex.
We look forward to describing follow-ups on these and other projects in future versions of the VSS annual conference.



