Our lab members are in Washington, DC this week to present their latest work on active vision, with a focus on how the primary visual cortex (V1) relates to the midbrain superior colliculus (SC) when it comes to generating rapid saccadic eye movements.
Comparison of the physiological properties of visual responses in primary visual cortex (V1) and superior colliculus (SC) neurons
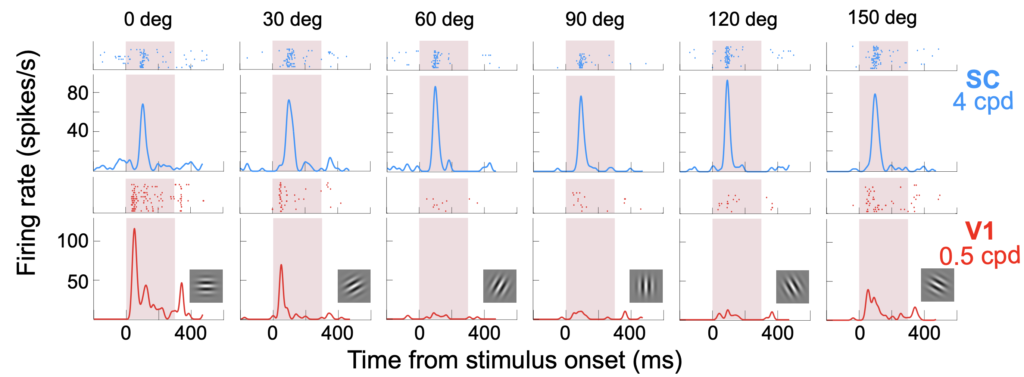
First off, Yue Yu describes her extensive comparisons of V1 and SC visual responses using paired recordings in the two brain areas. Among her many interesting findings are striking differences in the orientation tuning properties of SC and V1 neurons. SC neurons also show anisotropies in orientation selectivity as a function of preferred orientation, which V1 neurons do not exhibit.
Yue’s results, especially those on the timing of visual bursts in the two brain areas, have implications on our understanding of how V1 and SC contribute to eye movement generation.
V1 is both sufficient and necessary for saccadic inhibition, but other visual-motor pathways may become relevant with long-term V1 lesions
Next, Tanya looks more closely at how exogenous visual inputs are coordinated with internal saccade motor planning rhythms to optimally organize the timing and directions of eye movement behaviors. As is known, exogenous visual inputs cause a short-latency inhibition of saccade generation. However, how this inhibition happens exactly remains to be unknown. We previously speculated on the potential contributions of multiple brain areas, including V1, for this phenomenon. In her work, Tanya explicitly looks at V1 in particular, and using a variety of techniques, including electrical microstimulation, reversible inactivation, and permanent lesioning of V1.
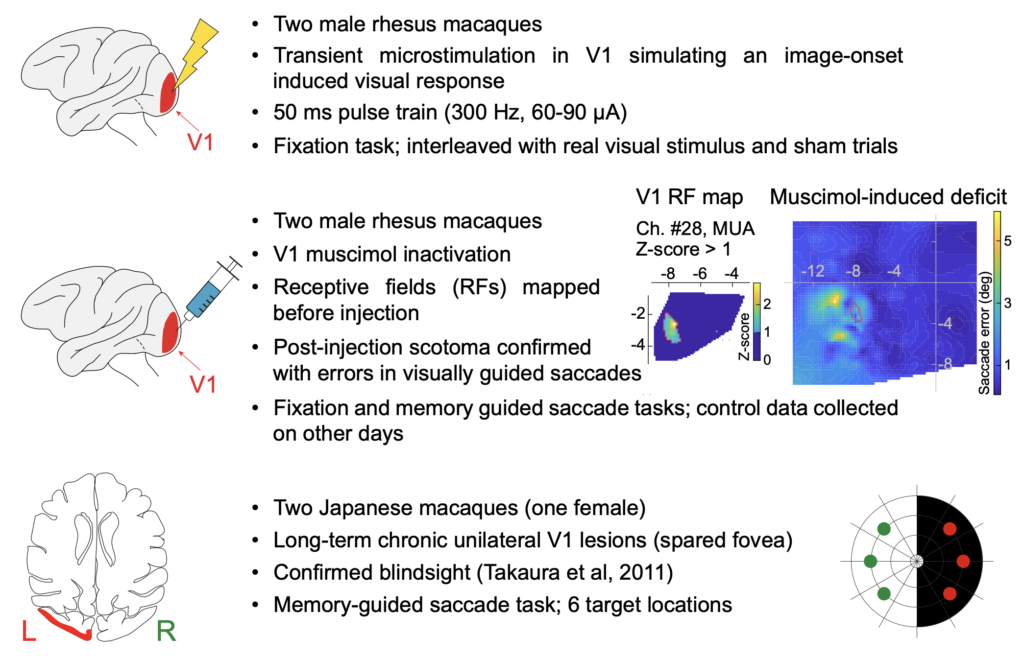
Tanya finds that transient focal inactivation of V1 leads to a complete abolition of saccadic inhibition. This is dramatically different from inactivation of the frontal cortex or the SC, inactivation of which does not alter saccadic inhibition in any meaningful way.
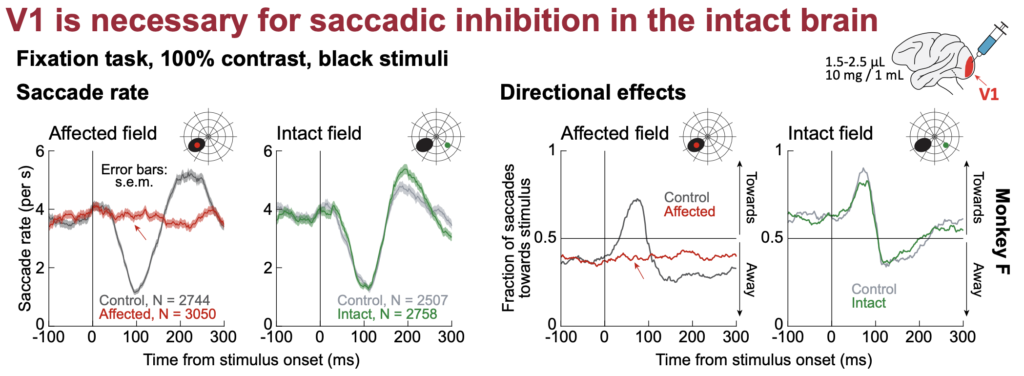
Interestingly, with a permanent V1 lesion instead of transient, temporary inactivation, saccadic inhibition re-emerges again, albeit with slightly different temporal dynamics. This suggests that other visual pathways from the retina to the oculomotor system, which bypass V1, may still be relevant for saccadic inhibition under certain circumstances.
Stay tuned for very exciting follow up work from Tanya directly exploring this intriguing question!
Both V1 and SC experience saccadic suppression, but with the SC being more in line with certain perceptual properties of saccadic suppression
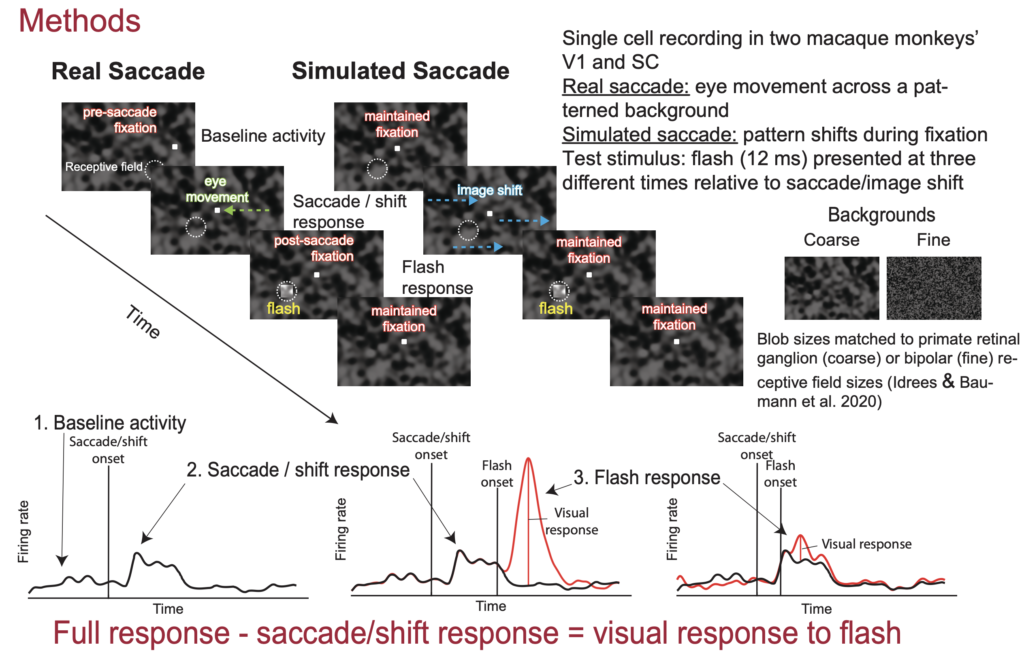
Continuing with the theme of active vision in V1 and SC, our third presentation at the conference is by Matthias Baumann. In his earlier work, like here and here, Matthias and Saad Idrees extensively studied the perceptual phenomenon of saccadic suppression. In this phenomenon, brief visual probes presented around the time of saccades are perceptually suppressed. Matthias and Saad found that this suppression remarkably starts in the retina, and they also found that the image-dependence of perceptual saccadic suppression reflects the image processing properties of retinal circuits. This leaves open the question of what happens downstream of the retina.
In his newest work, Matthias is recording from both V1 and SC neurons when saccades are generated across textured backgrounds. He also looks at the situation in which we simulate saccades by just shifting images rapidly over the retina without any “real” saccade generation by the brain. Matthias finds that both V1 and SC neurons experience “suppression” of visual sensitivity by saccades, whether real or simulated. Moreover, there are interesting dependencies of the suppression on the background texture and whether real or simulated saccades are generated. Intriguingly, V1 does not really behave like retinal circuits or perceptions. SC modulations are, on the other hand, a bit closer to the patterns of observations with perceptual saccadic suppression. These results generate new questions related to our understanding of the phenomenon of perceptual saccadic suppression.
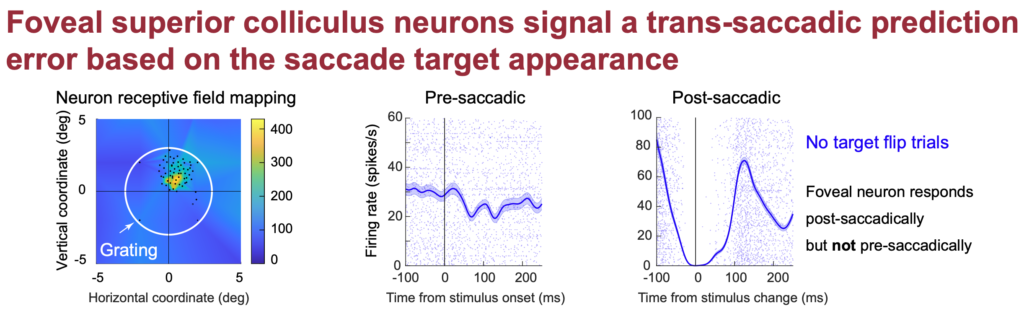
Foveal SC neurons anticipate the visual appearance of peripheral saccade targets in a predictive manner
Last, but not least, Tong looks at how the saccade motor program of the SC itself can help bridge perception across saccade. In recent work, like here and here, we showed how SC motor commands for saccades reflect the visual appearance of the peripheral saccade target. This raises the possibility that this information is available to make a prediction by the visual system about what the fovea will land on by the saccade. If so, then changing the visual appearance of the saccade target in mid-flight, when vision is most compromised, should influence how foveal neurons respond after the saccade. That is, the visual information impinging on these foveal neurons will be different from the “prediction” based on the peripheral saccade target appearance before the eye movement, and these foveal neurons might then signal this discrepancy in their activity. Tong shows exactly this phenomenon. Interestingly, when the peripheral appearance of the target is hard to distinguish, the predictive signal is also weak. Tong’s future work will attempt to understand the processes that ultimately jumpstart such a prediction process when the decision to make the eye movement is committed to, so stay tuned for exciting new work from Tong.